
Diversity of Harbinger-like Transposons in Teleost Fish Genomes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Genomes
2.2. TE Annotation
2.3. TE Genome Masking, Copy Number and Genome Coverage
2.4. Sequence Alignment and Phylogenetic Analysis
2.5. TE Expression Analysis in Spotted Gar, Zebrafish, Cod and Medaka
2.6. TE Expression Analysis in Medaka Gonads
2.7. TE Distribution on Medaka Chromosomes
3. Results
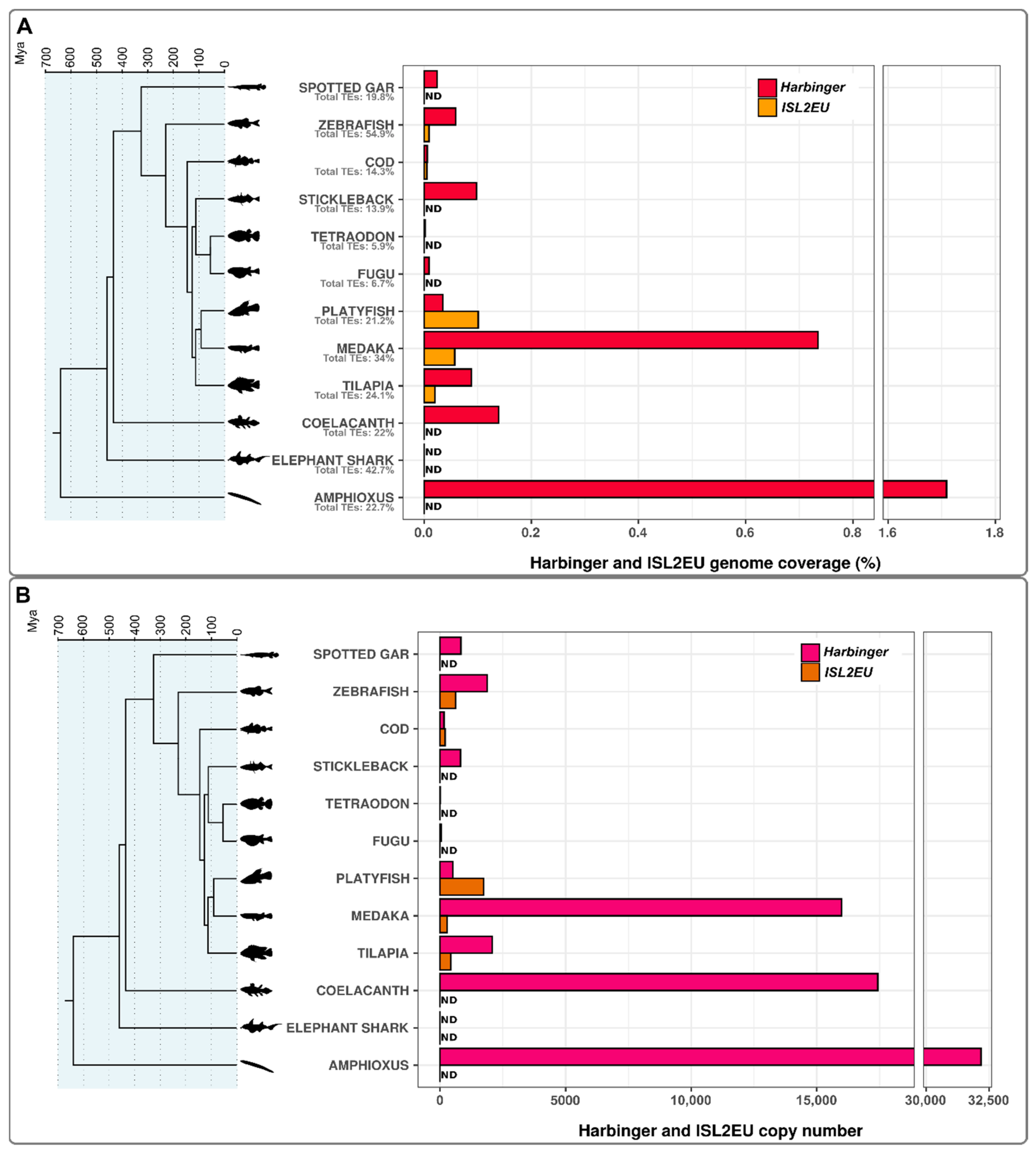
3.1. Differential Contributions of Harbinger-like Transposons to Fish Genomes
3.2. Distribution of Harbinger-like Transposons in Medaka Genome
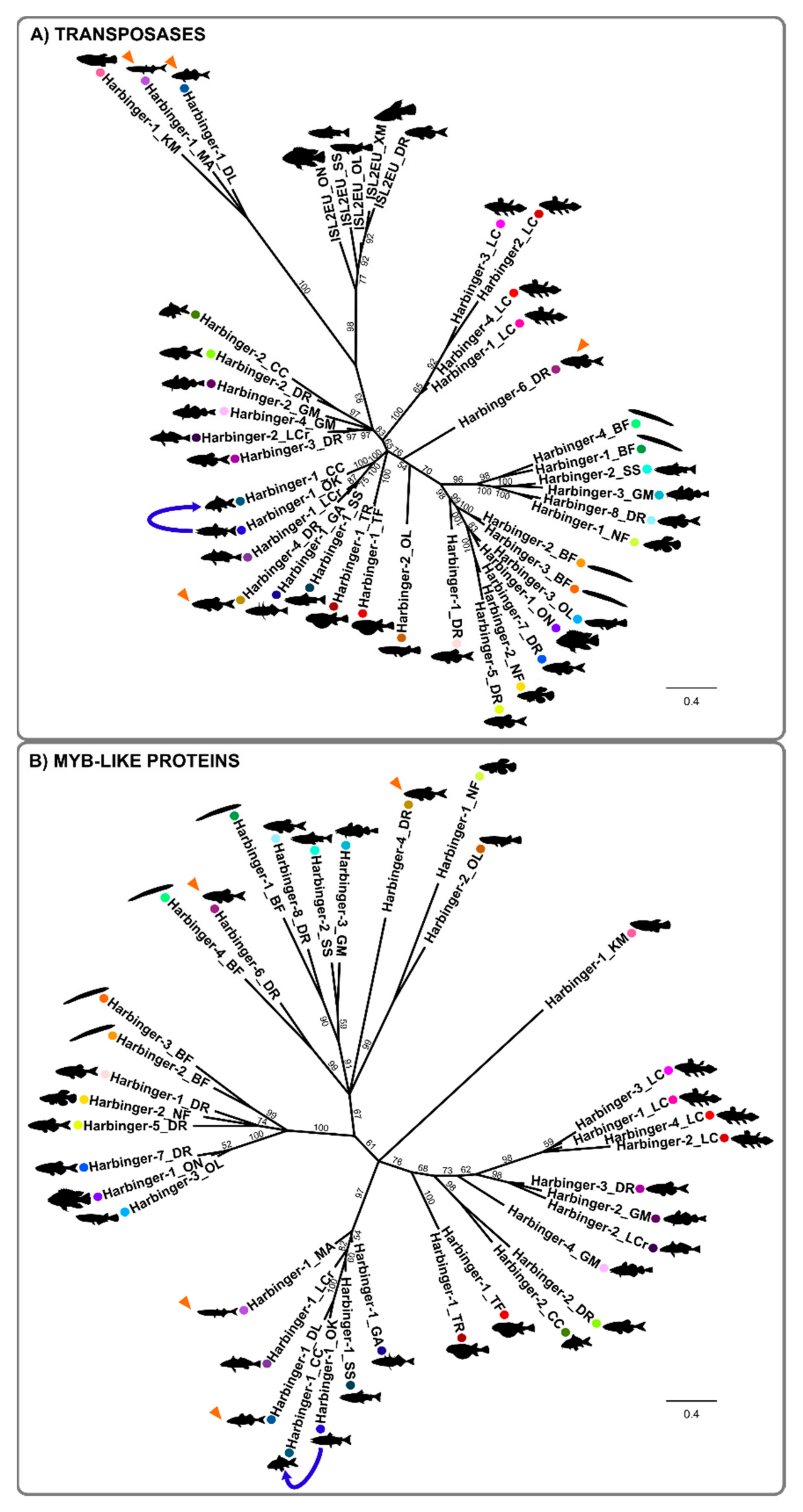
3.3. Evolution of Harbinger Transposons in Teleost Fish Genomes
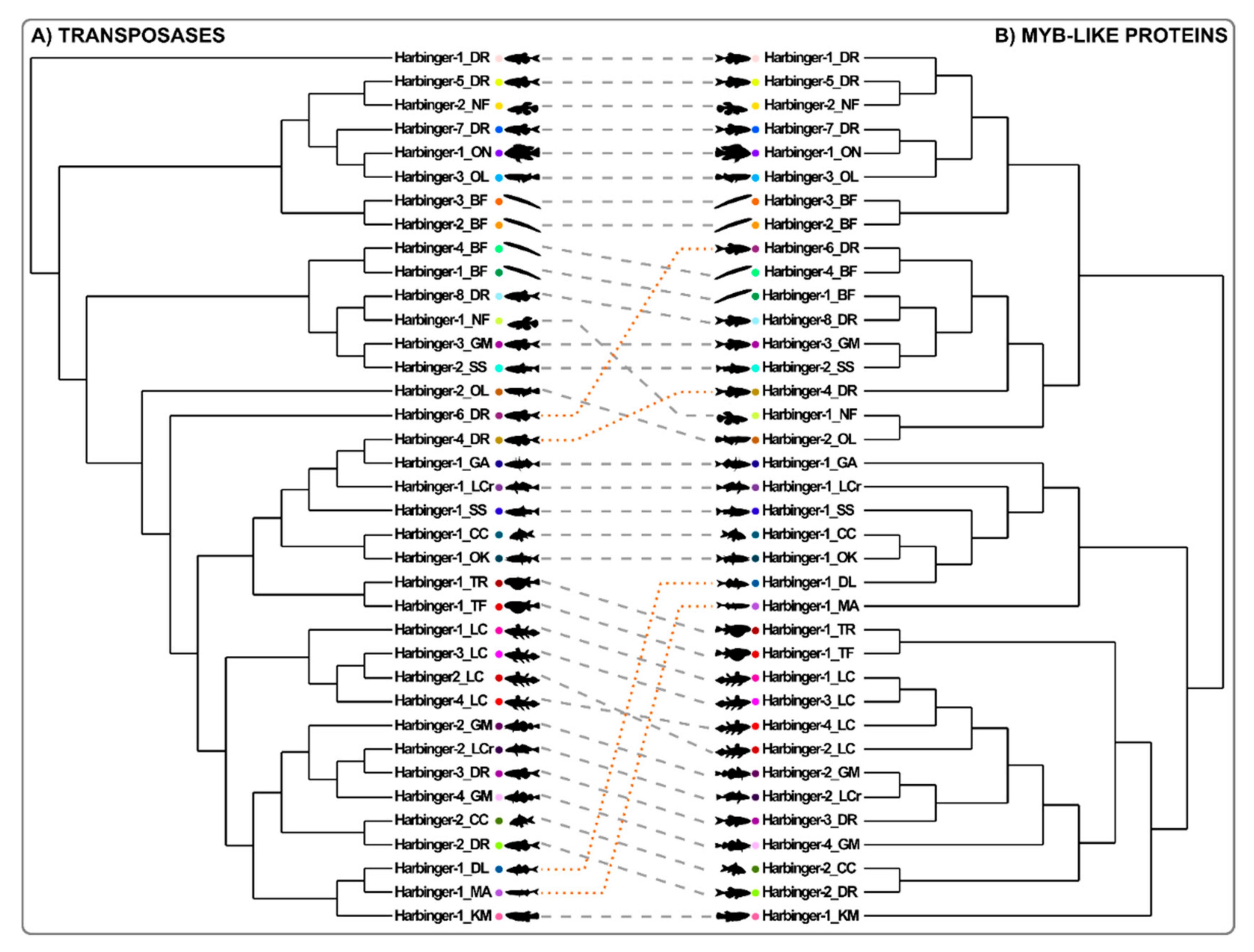
3.4. The Evolution of the Myb-like Proteins Recapitulates the Evolution of Transposase Proteins of Harbinger Transposons

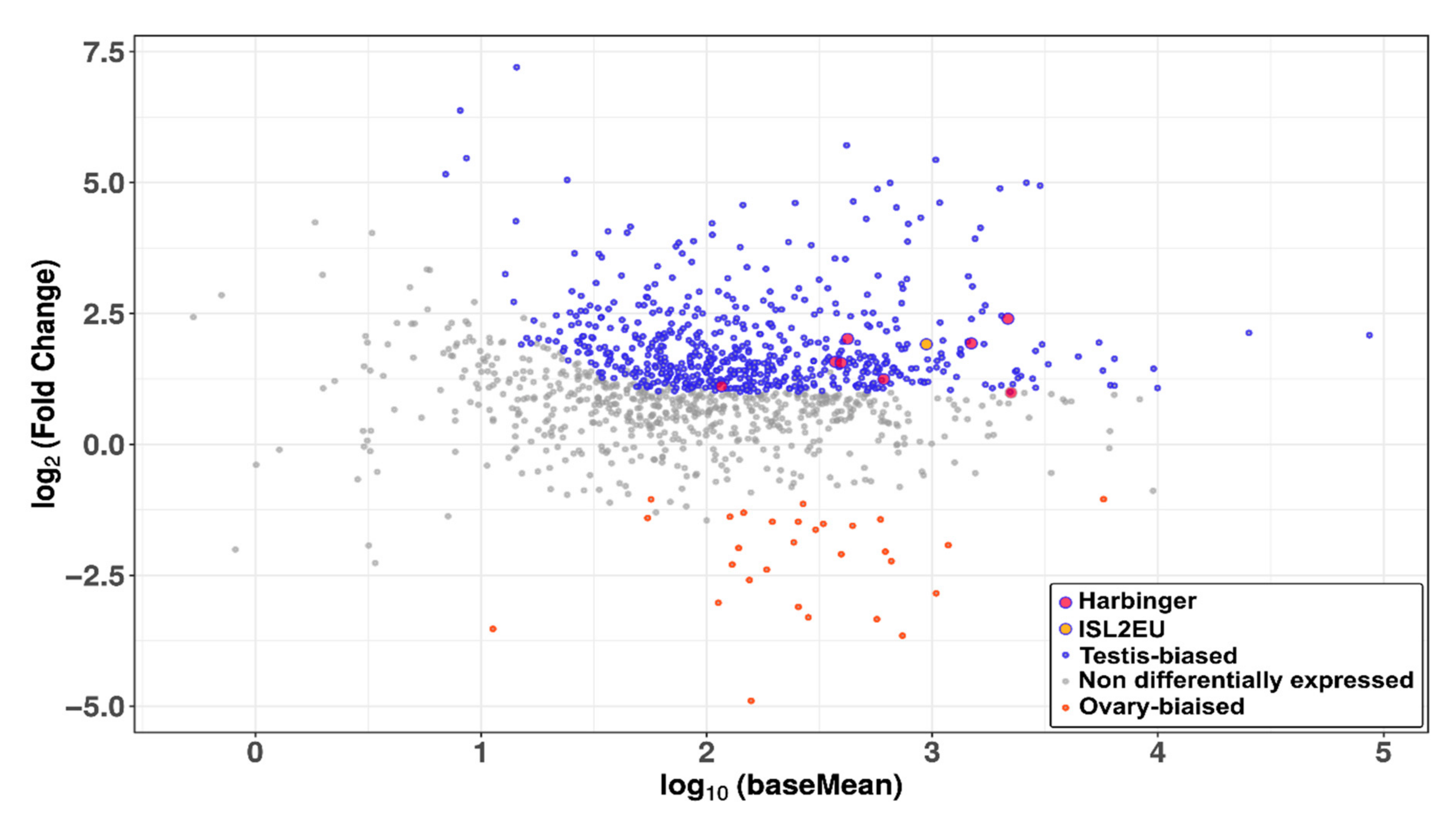
3.5. Harbinger-like Transposons Are Expressed in Fish
4. Discussion
4.1. Harbinger-like Transposons Are Inequitably Widespread in Fish Genomes
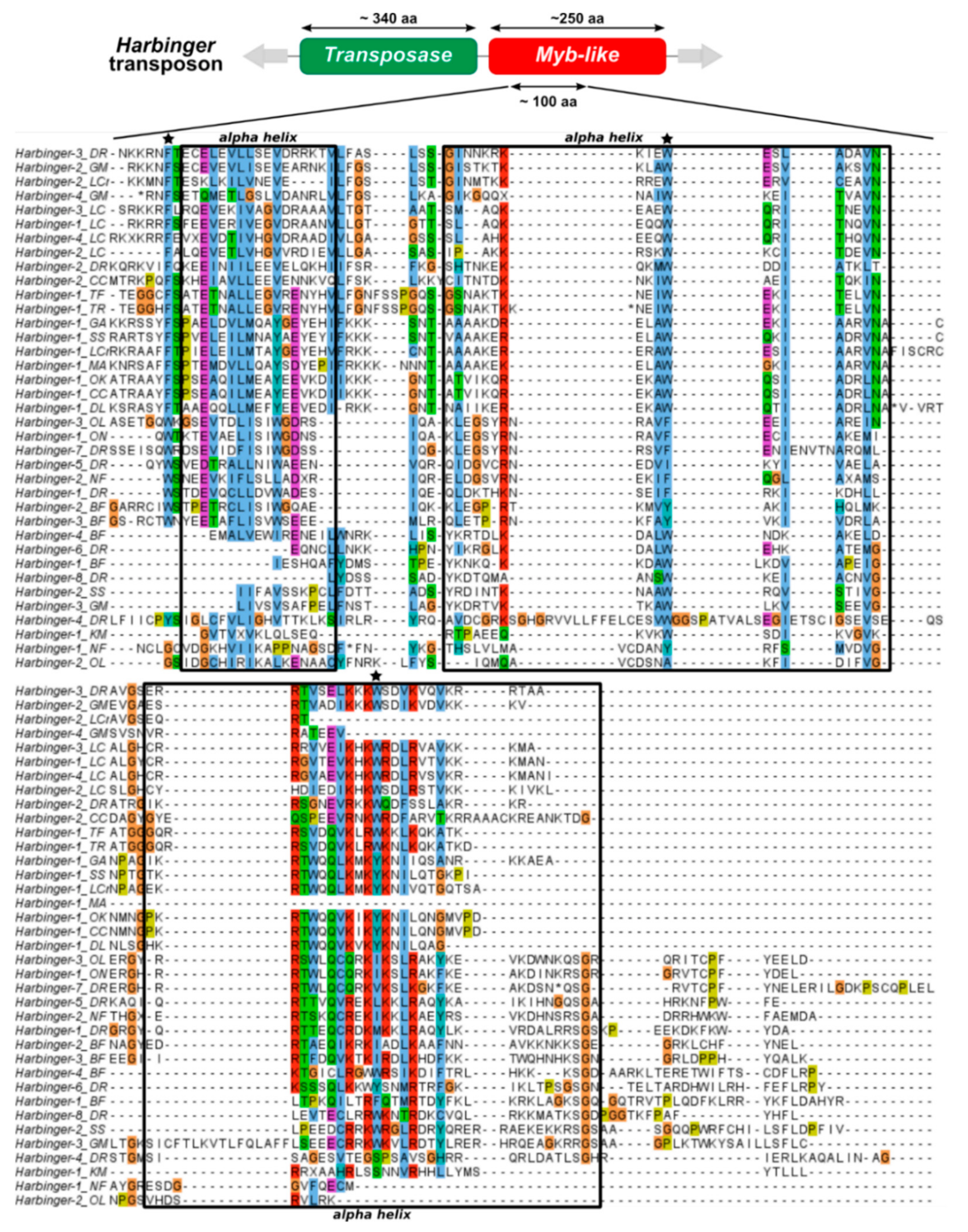
4.2. Evolutionary Relationships between the Two ORFs of Harbinger Transposons
4.3. Harbinger-Like Transposons Are Transcriptionally Active in Teleost Fish
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A Unified Classification System for Eukaryotic Transposable Elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Kidwell, M.G.; Lisch, D.R. Transposable Elements and Host Genome Evolution. Trends Ecol. Evol. 2000, 15, 95–99. [Google Scholar] [CrossRef]
- Volff, J.-N. Genome evolution and biodiversity in teleost fish. Heredity 2005, 94, 280–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory evolution of innate immunity through co-option of endogenous retroviruses. Science 2016, 351, 1083–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuong, E.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2016, 18, 71–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jangam, D.; Feschotte, C.; Betrán, E. Transposable Element Domestication as an Adaptation to Evolutionary Conflicts. Trends Genet. 2017, 33, 817–831. [Google Scholar] [CrossRef]
- Van Der Laan, R.; Eschmeyer, W.N.; Fricke, R. Family-group names of Recent fishes. Zootaxa 2014, 3882, 1–230. [Google Scholar] [CrossRef] [Green Version]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References; Electronic Version; The California Academy of Sciences: San Francisco, CA, USA, 2022. [Google Scholar]
- Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.-N. Comparative Analysis of Transposable Elements Highlights Mobilome Diversity and Evolution in Vertebrates. Genome Biol. Evol. 2015, 7, 567–580. [Google Scholar] [CrossRef]
- Carducci, F.; Barucca, M.; Canapa, A.; Carotti, E.; Biscotti, M.A. Mobile Elements in Ray-Finned Fish Genomes. Life 2020, 10, 221. [Google Scholar] [CrossRef]
- Kapitonov, V.V.; Jurka, J. Molecular Paleontology of Transposable Elements from Arabidopsis Thaliana. Genetica 1999, 107, 27–37. [Google Scholar] [CrossRef]
- Jurka, J.; Kapitonov, V.V. PIFs Meet Tourists and Harbingers: A Superfamily Reunion. Proc. Natl. Acad. Sci. USA 2001, 98, 12315–12316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Feschotte, C.; Zhang, Q.; Jiang, N.; Eggleston, W.B.; Wessler, S.R. P Instability Factor: An Active Maize Transposon System Associated with the Amplification of Tourist-like MITEs and a New Superfamily of Transposases. Proc. Natl. Acad. Sci. USA 2001, 98, 12572–12577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.; Bao, Z.; Zhang, X.; Hirochika, H.; Eddy, S.; McCouch, S.R.; Wessler, S.R. An active DNA transposon family in rice. Nature 2003, 421, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Terauchi, K.; Wada, M.; Hirano, H.-Y. The plant MITE mPing is mobilized in anther culture. Nature 2003, 421, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Kapitonov, V.V.; Jurka, J. Harbinger Transposons and an Ancient HARBI1 Gene Derived from a Transposase. DNA Cell Biol. 2004, 23, 311–324. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, N.; Feschotte, C.; Wessler, S.R. PIF- and Pong-Like Transposable Elements: Distribution, Evolution and Relationship with Tourist-Like Miniature Inverted-Repeat Transposable Elements. Genetics 2004, 166, 971–986. [Google Scholar] [CrossRef]
- Casola, C.; Lawing, A.M.; Betrán, E.; Feschotte, C. PIF-like Transposons are Common in Drosophila and Have Been Repeatedly Domesticated to Generate New Host Genes. Mol. Biol. Evol. 2007, 24, 1872–1888. [Google Scholar] [CrossRef] [Green Version]
- Grzebelus, D.; Lasota, S.; Gambin, T.; Kucherov, G.; Gambin, A. Diversity and structure of PIF/Harbinger-like elements in the genome of Medicago truncatula. BMC Genom. 2007, 8, 409. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.-W.; Wessler, S.R. The catalytic domain of all eukaryotic cut-and-paste transposase superfamilies. Proc. Natl. Acad. Sci. USA 2011, 108, 7884–7889. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.F.; Almeida, A.P.M.M.; Cota, J.; Pamphile, J.A.; da Silva, G.F.; de Araújo, E.F.; Gramacho, K.P.; Brommonschenkel, S.H.; Pereira, G.A.G.; de Queiroz, M.V. Boto, a class II transposon in Moniliophthora perniciosa, is the first representative of the PIF/Harbinger superfamily in a phytopathogenic fungus. Microbiology 2013, 159, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Han, M.-J.; Xiong, C.-L.; Zhang, H.-B.; Zhang, M.-Q.; Zhang, H.-H.; Zhang, Z. The diversification of PHIS transposon superfamily in eukaryotes. Mob. DNA 2015, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Markova, D.N.; Mason-Gamer, R.J. Diversity, abundance, and evolutionary dynamics of Pong-like transposable elements in Triticeae. Mol. Phylogenetics Evol. 2015, 93, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinzelle, L.; Kapitonov, V.V.; Grzela, D.P.; Jursch, T.; Jurka, J.; Izsvák, Z.; Ivics, Z. Transposition of a Reconstructed Harbinger Element in Human Cells and Functional Homology with Two Transposon-Derived Cellular Genes. Proc. Natl. Acad. Sci. USA 2008, 105, 4715–4720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.-J.; Xu, H.-E.; Zhang, H.-H.; Feschotte, C.; Zhang, Z. Spy: A New Group of Eukaryotic DNA Transposons without Target Site Duplications. Genome Biol. Evol. 2014, 6, 1748–1757. [Google Scholar] [CrossRef] [Green Version]
- Hancock, C.N.; Zhang, F.; Wessler, S.R. Transposition of the Tourist-MITE mPing in yeast: An assay that retains key features of catalysis by the class 2 PIF/Harbinger superfamily. Mob. DNA 2010, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elewa, A.; Wang, H.; Talavera-López, C.; Joven, A.; Brito, G.; Kumar, A.; Hameed, L.S.; Penrad-Mobayed, M.; Yao, Z.; Zamani, N.; et al. Reading and editing the Pleurodeles waltl genome reveals novel features of tetrapod regeneration. Nat. Commun. 2017, 8, 2286. [Google Scholar] [CrossRef]
- Markova, D.N.; Mason-Gamer, R.J. Transcriptional Activity of PIF and Pong-like Class II Transposable Elements in Triticeae. BMC Evol. Biol. 2017, 17, 178. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.J.; Sumiyama, K.; Amemiya, C.T. A Living Fossil in the Genome of a Living Fossil: Harbinger Transposons in the Coelacanth Genome. Mol. Biol. Evol. 2012, 29, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Kapitonov, V.V.; Jurka, J. A Universal Classification of Eukaryotic Transposable Elements Implemented in Repbase. Nat. Rev. Genet. 2008, 9, 411–412, author reply 414. [Google Scholar] [CrossRef]
- Amemiya, C.T.; Alföldi, J.; Lee, A.P.; Fan, S.; Philippe, H.; Maccallum, I.; Braasch, I.; Manousaki, T.; Schneider, I.; Rohner, N.; et al. The African Coelacanth Genome Provides Insights into Tetrapod Evolution. Nature 2013, 496, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast Selection of Best-Fit Models of Protein Evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [Green Version]
- Pasquier, J.; Cabau, C.; Nguyen, T.; Jouanno, E.; Severac, D.; Braasch, I.; Journot, L.; Pontarotti, P.; Klopp, C.; Postlethwait, J.H.; et al. Gene Evolution and Gene Expression after Whole Genome Duplication in Fish: The PhyloFish Database. BMC Genom. 2016, 17, 368. [Google Scholar] [CrossRef] [Green Version]
- Mariette, J.; Noirot, C.; Nabihoudine, I.; Bardou, P.; Hoede, C.; Djari, A.; Cabau, C.; Klopp, C. RNAbrowse: RNA-Seq de Novo Assembly Results Browser. PLoS ONE 2014, 9, e96821. [Google Scholar] [CrossRef] [Green Version]
- Dechaud, C.; Miyake, S.; Martinez-Bengochea, A.; Schartl, M.; Volff, J.-N.; Naville, M. Clustering of Sex-Biased Genes and Transposable Elements in the Genome of the Medaka Fish Oryzias Latipes. Genome Biol. Evol. 2021, 13, evab230. [Google Scholar] [CrossRef]
- Yang, W.R.; Ardeljan, D.; Pacyna, C.N.; Payer, L.M.; Burns, K.H. SQuIRE Reveals Locus-Specific Regulation of Interspersed Repeat Expression. Nucleic Acids Res. 2019, 47, e27. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Peccoud, J.; Xu, M.-R.-X.; Zhang, X.-G.; Gilbert, C. Horizontal Transfer and Evolution of Transposable Elements in Vertebrates. Nat. Commun. 2020, 11, 1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vienne, D.M.; Giraud, T.; Martin, O.C. A Congruence Index for Testing Topological Similarity between Trees. Bioinformatics 2007, 23, 3119–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galbraith, J.D.; Ludington, A.J.; Sanders, K.L.; Suh, A.; Adelson, D.L. Horizontal Transfer and Subsequent Explosive Expansion of a DNA Transposon in Sea Kraits (Laticauda). Biol. Lett. 2021, 17, 20210342. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Wang, Z.-J.; Li, Q.-Y.; Lian, J.-M.; Zhou, Y.; Lu, B.-Z.; Jin, L.-J.; Qiu, P.-X.; Zhang, P.; Zhu, W.-B.; et al. Evolutionary Trajectories of Snake Genes and Genomes Revealed by Comparative Analyses of Five-Pacer Viper. Nat. Commun. 2016, 7, 13107. [Google Scholar] [CrossRef] [PubMed]
- Stow, E.C.; Kaul, T.; deHaro, D.L.; Dem, M.R.; Beletsky, A.G.; Morales, M.E.; Du, Q.; LaRosa, A.J.; Yang, H.; Smither, E.; et al. Organ-, Sex- and Age-Dependent Patterns of Endogenous L1 MRNA Expression at a Single Locus Resolution. Nucleic Acids Res. 2021, 49, 5813–5831. [Google Scholar] [CrossRef]
- Zempleni, J.; Chew, Y.C.; Bao, B.; Pestinger, V.; Wijeratne, S.S.K. Repression of Transposable Elements by Histone Biotinylation. J. Nutr. 2009, 139, 2389–2392. [Google Scholar] [CrossRef] [Green Version]
- Rebollo, R.; Romanish, M.T.; Mager, D.L. Transposable Elements: An Abundant and Natural Source of Regulatory Sequences for Host Genes. Annu. Rev. Genet. 2012, 46, 21–42. [Google Scholar] [CrossRef]
- Iwasaki, Y.W.; Siomi, M.C.; Siomi, H. PIWI-Interacting RNA: Its Biogenesis and Functions. Annu. Rev. Biochem. 2015, 84, 405–433. [Google Scholar] [CrossRef]
- Sarkar, A.; Volff, J.-N.; Vaury, C. PiRNAs and Their Diverse Roles: A Transposable Element-Driven Tactic for Gene Regulation? FASEB J. 2017, 31, 436–446. [Google Scholar] [CrossRef] [Green Version]
- Kneitz, S.; Mishra, R.R.; Chalopin, D.; Postlethwait, J.; Warren, W.C.; Walter, R.B.; Schartl, M. Germ Cell and Tumor Associated PiRNAs in the Medaka and Xiphophorus Melanoma Models. BMC Genom. 2016, 17, 357. [Google Scholar] [CrossRef] [Green Version]
- Etchegaray, E.; Naville, M.; Volff, J.-N.; Haftek-Terreau, Z. Transposable Element-Derived Sequences in Vertebrate Development. Mob DNA 2021, 12, 1. [Google Scholar] [CrossRef]
- Cosby, R.L.; Chang, N.-C.; Feschotte, C. Host-Transposon Interactions: Conflict, Cooperation, and Cooption. Genes Dev 2019, 33, 1098–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.C.; Hartwig, B.; Perera, P.; Mora-García, S.; de Leau, E.; Thornton, H.; de Lima Alves, F.; de Alves, F.L.; Rappsilber, J.; Rapsilber, J.; et al. Kicking against the PRCs—A Domesticated Transposase Antagonises Silencing Mediated by Polycomb Group Proteins and Is an Accessory Component of Polycomb Repressive Complex 2. PLoS Genet. 2015, 11, e1005660. [Google Scholar] [CrossRef]
- Duan, C.-G.; Wang, X.; Xie, S.; Pan, L.; Miki, D.; Tang, K.; Hsu, C.-C.; Lei, M.; Zhong, Y.; Hou, Y.-J.; et al. A Pair of Transposon-Derived Proteins Function in a Histone Acetyltransferase Complex for Active DNA Demethylation. Cell Res. 2017, 27, 226–240. [Google Scholar] [CrossRef]
- Velanis, C.N.; Perera, P.; Thomson, B.; de Leau, E.; Liang, S.C.; Hartwig, B.; Förderer, A.; Thornton, H.; Arede, P.; Chen, J.; et al. The Domesticated Transposase ALP2 Mediates Formation of a Novel Polycomb Protein Complex by Direct Interaction with MSI1, a Core Subunit of Polycomb Repressive Complex 2 (PRC2). PLoS Genet. 2020, 16, e1008681. [Google Scholar] [CrossRef] [PubMed]
- Cosby, R.L.; Judd, J.; Zhang, R.; Zhong, A.; Garry, N.; Pritham, E.J.; Feschotte, C. Recurrent Evolution of Vertebrate Transcription Factors by Transposase Capture. Science 2021, 371, eabc6405. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; He, J.; Velanis, C.N.; Zhu, Y.; He, Y.; Tang, K.; Zhu, M.; Graser, L.; de Leau, E.; Wang, X.; et al. A Domesticated Harbinger Transposase Forms a Complex with HDA6 and Promotes Histone H3 Deacetylation at Genes but Not TEs in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 1462–1474. [Google Scholar] [CrossRef] [PubMed]
- Deneweth, J.; Van de Peer, Y.; Vermeirssen, V. Nearby Transposable Elements Impact Plant Stress Gene Regulatory Networks: A Meta-Analysis in A. Thaliana and S. Lycopersicum. BMC Genom. 2022, 23, 18. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Etchegaray, E.; Dechaud, C.; Barbier, J.; Naville, M.; Volff, J.-N. Diversity of Harbinger-like Transposons in Teleost Fish Genomes. Animals 2022, 12, 1429. https://doi.org/10.3390/ani12111429
Etchegaray E, Dechaud C, Barbier J, Naville M, Volff J-N. Diversity of Harbinger-like Transposons in Teleost Fish Genomes. Animals. 2022; 12(11):1429. https://doi.org/10.3390/ani12111429
Chicago/Turabian StyleEtchegaray, Ema, Corentin Dechaud, Jérémy Barbier, Magali Naville, and Jean-Nicolas Volff. 2022. "Diversity of Harbinger-like Transposons in Teleost Fish Genomes" Animals 12, no. 11: 1429. https://doi.org/10.3390/ani12111429
APA StyleEtchegaray, E., Dechaud, C., Barbier, J., Naville, M., & Volff, J.-N. (2022). Diversity of Harbinger-like Transposons in Teleost Fish Genomes. Animals, 12(11), 1429. https://doi.org/10.3390/ani12111429